
سیتوژنتیک شاخهای از ژنتیک است که ساختار DNA را در هسته سلول بررسی میکند. DNA در حین تقسیم سلولی متراکم میشود و کروموزومها را تشکیل میدهد. سیتوژنتیک تعداد و مورفولوژی کروموزومها را بررسی میکند. با استفاده از تکنیکهای باندینگ کروموزوم (سیتوژنتیک کلاسیک) یا هیبریداسیون پروبهای دارای برچسب فلورسانس (سیتوژنتیک مولکولی) بررسی کروموزومها در مطالعات سیتوژنتیک قابل انجام است.
تعداد و مورفولوژی کروموزومهای موجود در سلول یک گونه خاص همیشه در اکثر سلولهای بدن (به استثنای سلولهای تولید مثلی و سایر موارد مانند کبد) ثابت است. این ویژگی در هر گونه منحصر به فرد است، به عنوان مثال در انسان تعداد کروموزومها برابر با ۴۶ عدد است.
سیتوژنتیک چیست؟
همان طور که در بالا اشاره شد، سیتوژنتیک در اصل بخش مهمی از مطالعات ژنتیکی را در بر میگیرد، از سویی دیگر این شاخه از ژنتیک بخشی از زیست شناسی سلولی یا سیتولوژی (زیرمجموعه آناتومی انسانی) به شمار میآید که مربوط به چگونگی ارتباط کروموزومها با رفتار سلولی، به ویژه چگونگی رفتار سلولها در هنگام میتوز و میوز است.
تکنیکهای مورد استفاده در مطالعات سیتوژنتیک شامل کاریوتایپینگ، تجزیه و تحلیل کروموزومهای باند G، سایر روشهای باند سیتوژنتیک و همچنین سیتوژنتیک مولکولی مانند هیبریداسیون درجا (FISH) و هیبریداسیون ژنومی مقایسهای (CGH) است.
زیست شناسی مولکولی و سیتوژنتیک نقش مهمی در تشخیص ناهنجاریهای کروموزومی مرتبط با سرطان دارند و همچنین در توصیف ناهنجاریهای جدیدی که ممکن است باعث بروز بیماری شوند، مورد استفاده قرار میگیرند.
باندینگ کروموزوم
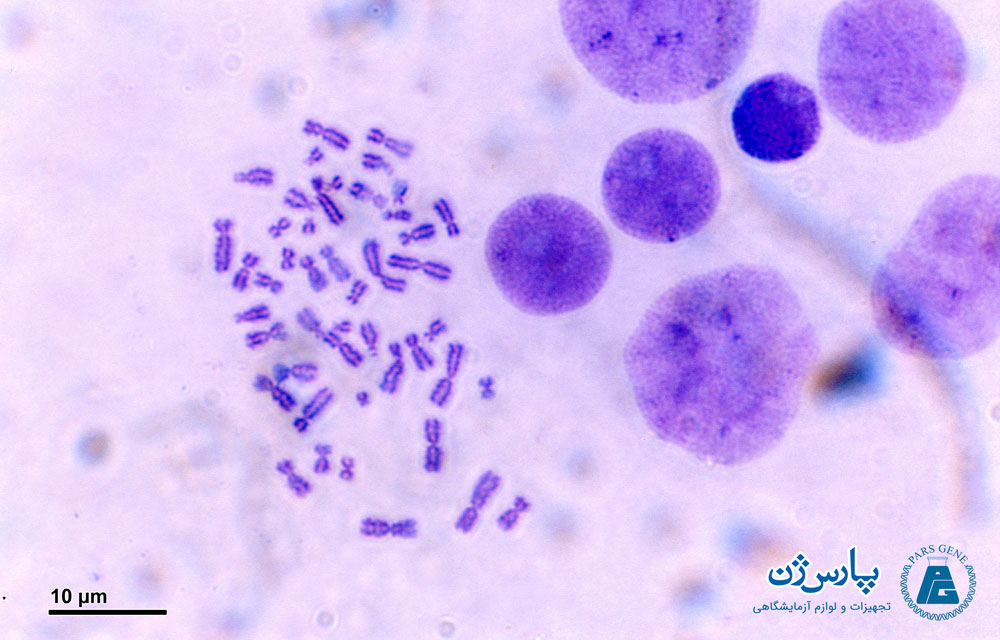
کروموزومهای متافاز را میتوان با استفاده از تکنیکهای خاص رنگ آمیزی، به اصطلاح «باندینگ» (Banding) شناسایی کرد. برای این کار، سلولها کشت میشوند و در مرحله متافاز تقسیم سلولی متوقف میشوند تا تعداد سلولهای مناسب برای انجام باندینگ بیشتر شود. این سلولها سپس روی یک اسلاید میکروسکوپی پخش میشوند، با رنگ مناسب آغشته شده و با استفاده از میکروسکوپ مورد بررسی قرار میگیرند. اکثر آنالیزهای سیتوژنتیک متداول، به کاریوتایپ باند کروموزومهای متافاز بستگی دارند.
یک باند به عنوان بخشی از کروموزوم تعریف شده است که با ظاهر تیرهتر یا روشنتر با یک یا چند روش باندینگ، کاملاً از بخشهای مجاور آن قابل تشخیص است. در این حالت کروموزومها به عنوان یک مجموعه از نوارهای روشن و تاریک متوالی قابل مشاهده میشوند.
انواع باندینگ کروموزوم
باندهای کروموزومی به مناطق روشن وتیره متناوب در طول یک کروموزوم اشاره دارد که پس از رنگ آمیزی با یک رنگ تولید می شوند. باند به عنوان بخشی از کروموزوم تعریف میشود که با ظاهر تیرهتر یا روشنتر با استفاده از یک یا چند روش باندینگ، کاملاً از بخشهای مجاور آن قابل تشخیص است. دانشمندی به نام «کاسپرسون» (Torbjorn Caspersson) هنگام کار با گیاهان و استفاده از خردل کیناکرین (Quinacrine) با فلورسنت بالا، اولین الگوهای باند کروموزومی را تولید کرد که در سال ۱۹۶۸ گزارش شد.
باند های متناوب فلورسانس روشن و تیره باند Q نامیده میشدند. باندهای کیناکرین روشن در درجه اول از DNA غنی از بازهای آدنین و تیمین تشکیل شده بودند. رنگ آمیزی خردل کیناکرین در سال ۱۹۷۰ در کروموزومهای انسانی به کار گرفته شد تا اولین باندهای کاریوگرام انسانی را بسازد. پیش از این مشاهده باندینگ، ۲۴ کروموزوم انسانی (۲۲ جفت اتوزومال و کروموزومهای جنسی X و Y) شناسایی شده و به هفت گروه جداگانه در گروههای مختلف (از A تا G) براساس اندازه و موقعیت سانترومر تقسیم شده بودند.
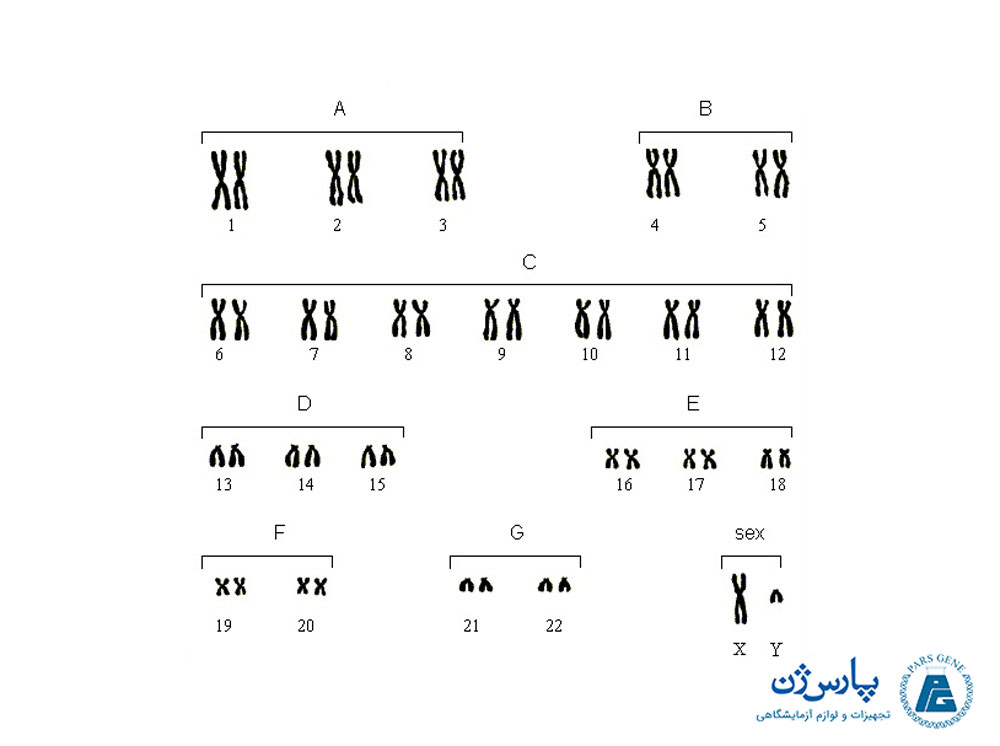
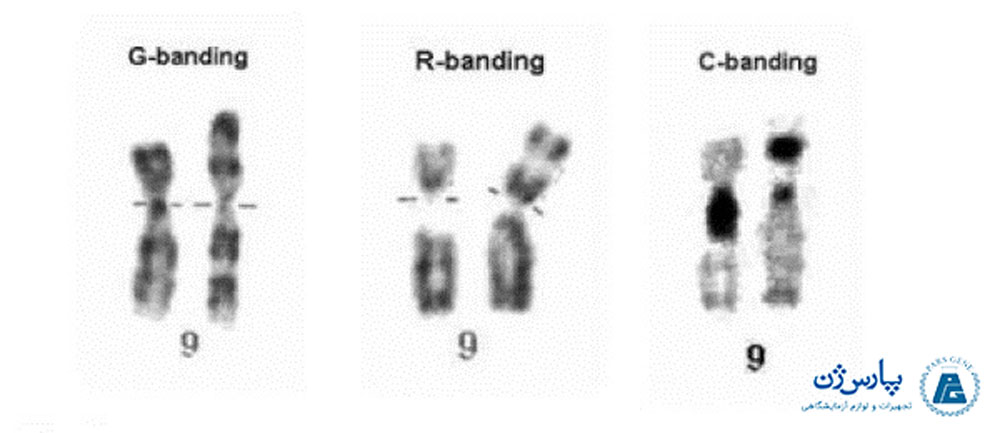
تجزیه و تحلیل کروموزومهای باند Q منجر به شناسایی هر یک از کروموزومهای انسانی و مناطق و گروههای خاص در درون هر کروموزوم شد. در سال ۱۹۷۶ کمیته دائمی بین المللی نامگذاری سیتوژنتیک انسانی ایجاد شد تا همچنان به بهبود نامگذاری کروموزوم بپردازد. بلافاصله بعد از معرفی باند Q، چندین روش رنگ آمیزی دیگر معرفی شدند که انواع مختلفی از الگوها و نوارها از جمله C-banding (که به طور متفاوتی سانترومر هر یک از کروموزومها را رنگ آمیزی میکرد)، Gباندینگ (G-banding) و R باندینگ ((R- banding تولید کردند.
اتصال باندینگ G معمولاً با استفاده از آنزیم پروتئولیتیک مانند تریپسین و به دنبال آن رنگآمیزی با روش گیمسا (Giemsa) که به DNA اتصال مییابد، صورت میگیرد. باندهای G یک الگوی باندینگ ایجاد میکنند که می تواند با باند Q در ارتباط باشد، به طوری که نوارهای «روشن G» (G-light) معادل مناطق «تیره Q» (Q-dull) و نوارهای «تیره G» (G-dark) معادل مناطق درخشان فلورسنت هستند. روش باندینگ G به دلیل ماندگاری و وضوح آن، به عنوان رایجترین روش رنگ آمیزی در سیتوژنتیک انسان به شمار میآید (در مقایسه با سیگنال فلورسانس محو، تولید شده توسط باند Q).
انواع تکنیکهای باندینگ
تکنیکهای باندینگ در دو گروه اصلی قرار میگیرند:
- روشهایی که منجر به ایجاد باند در طول کل کروموزوم میشوند، مانند باندهای G، Q و.R باندهای G و R میتوانند زمینه روشن یا فلورسنت باشند.
- روشهایی که تعداد محدودی از باندها یا ساختارهای خاص را رنگآمیزی میکنند. این گروه، روشهایی را در بر میگیرد که باندهای سانترومر، گروههای C و مناطق سازمان دهنده هستهای، NOR (در مناطق پایانی کروموزومهای آکروسانتریک) را نشان میدهند. روشهای باندینگ C اجازه شناسایی هر کروموزوم موجود در سلول سوماتیک را نمیدهد، اما میتواند برای شناسایی کروموزومهای خاص استفاده شود.
باند G زمینه روشن
این باندهای G بیشترین استفاده را دارند. آنها نام خود را از رنگ Giemsa میگیرند، اما میتوانند با رنگهای دیگر تولید شوند. در باندهای G، مناطق تاریک به صورت هتروکروماتین، با همانند سازی طولانی مدت و غنی از AT هستند. مناطق روشن اغلب از نوع یوکروماتین هستند، همانند سازی سریع دارند و دارای توالیهای غنی از GC هستند.
باندینگ G
باندینگ Gیا باند گیمسا Giemsa روشی است که در سیتوژنتیک برای تولید کاریوتایپ قابل مشاهده با رنگ آمیزی کروموزومهای متراکم مورد استفاده قرار میگیرد. روش گیمسا برای شناسایی بیماریهای ژنتیکی از طریق نمایش تصاویری از کل کروموزومهای یک گونه مفید است.

در روش G باندینگ کروموزومهای متافاز با تریپسین (برای هضم بخشی از کروموزوم) تیمار میشوند و با رنگ گیمسا مورد رنگ آمیزی قرار میگیرند. مناطق هتروکروماتیک که DNA آنها دارای توالیهای غنی شده از آدنین و تیمین و تعداد بسیار کمی ژن هستند، در روش باندینگG به شکل تیر رنگ آمیزی میشوند. در مقابل، کروموزومهای کمتر متراکم یا مناطق یوکروماتین که دارای توالیهای غنی گوانین و سیتوزین هستند و فعالیت بالایی از نظررونویسی دارند، کمتر رنگ گیمسا را جذب کرده و از این رو به صورت نوارهای روشنی در باندینگ G مشاهده میشوند. الگوی نوارها روی هر بازوی کروموزوم از سانترومر تا تلومر شماره گذاری میشود. این سیستم شماره گذاری اجازه میدهد تا هر باند روی کروموزوم دقیقاً مشخص و توصیف شود. معکوس باندهای G در باندینگ به روشR بدست میآید. از این باندها میتوان برای شناسایی ناهنجاریهای کروموزومی مانند «جابجایی» (Translocations) استفاده کرد، زیرا برای هر کروموزوم الگوی منحصر به فردی از باندهای روشن و تاریک وجود دارد.
شناسایی و گروهبندی کروموزومها براساس رنگ آمیزی، کار دشواری است، زیرا رنگ یکنواخت کروموزومها باعث میشود تمایز بین کروموزومهای مختلف دشوار شود. بنابراین، تکنیکی مانند باندینگ G ایجاد شده است که باعث میشود، باندها روی کروموزومها ظاهر شوند. این گروهها از نظر ظاهری در کروموزومهای همولوگ یکسان بودند، بنابراین با رنگ آمیزی شناسایی آسانتر و دقیقتر میشود. هرچه کروموزومها کمتر متراکم باشند، در هنگام رنگ آمیزی باندG ، باندهای بیشتری ظاهر میشوند. این بدان معنی است که کروموزومهای مختلف از نظر ظاهری در متافاز از پروفاز متمایز هستند.
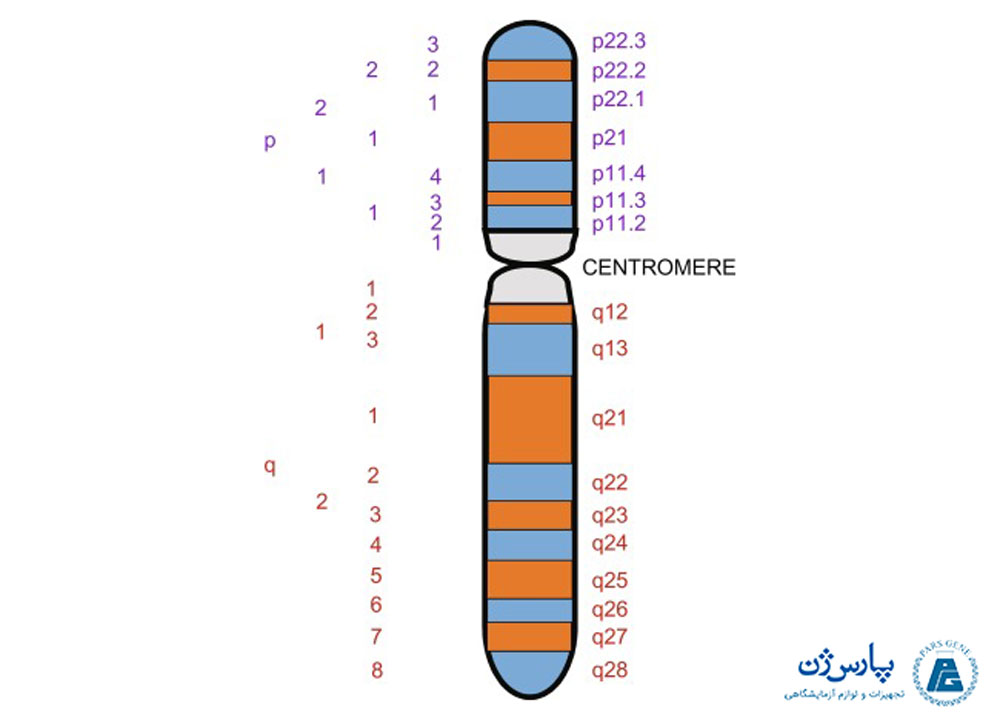
باند G اجازه میدهد تا هر کروموزوم با الگوی باند اختصاصی مشخص شود. الگوی باندینگ میتواند ناهنجاریهای کروموزومی یا بازآرایی ساختاری مانند جابجاییها، حذفها، جایگیری و وارونگیها را تشخیص دهد. باند G به مناطق، باندها و زیر باندهای مختلف تقسیم میشود. شکل زیر نمودار متداول از باندهای کروموزوم X انسان را نشان میدهد. به طور کلی، باندهایی که از نظر بازهای گوانین و سیتوزین نسبت به مناطق بین باندی فقیرتر هستند، نواحی را در بر دارند که ژنها در آنها جای میگیرند. تیمار با شرایط سخت برای کروموزوم (۸۷ درجه سانتیگراد به مدت ۱۰ دقیقه) قبل از رنگ آمیزی گیمسا، میتواند الگویی به نام باند R تولید کند که خلاف الگوی باندینگ G است. باند R میتواند ناحیه یوکروماتین را مورد رنگ آمیزی قرار دهد.
منطقهای که از مرکز سانترومر عبور میکند، غالباً مقدار قابل توجهی هتروکروماتین را نشان میدهد. وقتی کل کروموزوم متراکم شود، هتروکروماتین سانترومری در یک کروموزوم میتوزی قابل مشاهده نیست. با این وجود، سانترومر با رنگ آمیزی که باندهای C ایجاد میکنند، قابل مشاهده میشود. باند C یا باند سانترومر نتیجه تیمار قلیایی کروموزوم است. رنگ آمیزی سانترومر در الگوهای باند G وجود ندارد. باندهای C با هتروکروماتین در امتداد کروموزومها و اطراف سانترومرها همراه هستند.
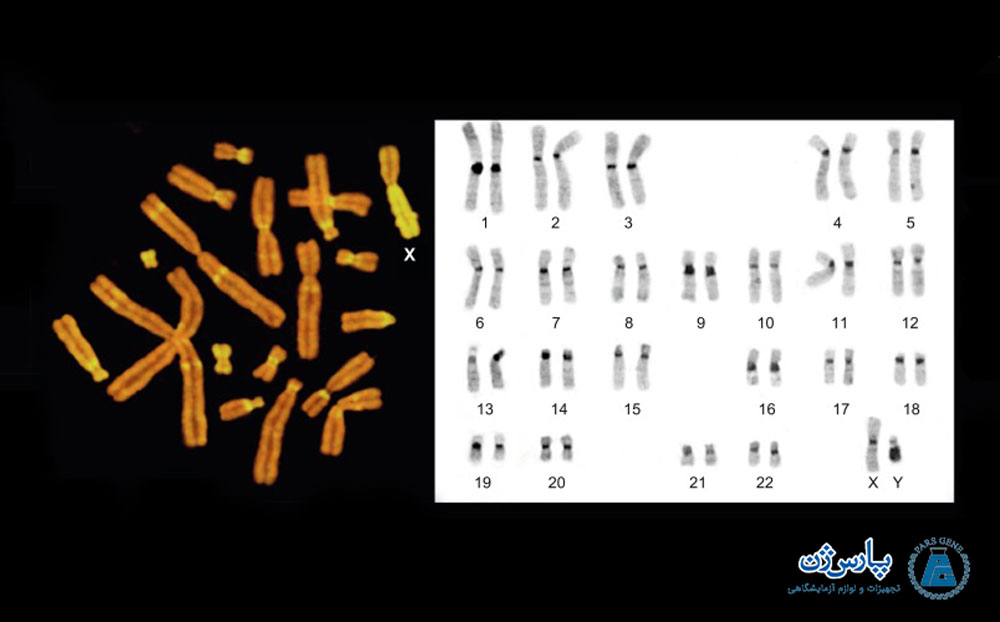
روشها رنگآمیزی از قبیل باندینگ C، رنگآمیزی نقرهای از مناطق سازمان دهنده هستهای، باندینگ T مناطق تلومر و رنگ آمیزی آنتی بادی از اجزای کروموزوم با باندهای کمتری مورد هدف قرار میگیرند و در سیتوژنتیک کاربرد چندانی ندارند.
چهار گروه از باندها قابل تشخیص هستند:
- باندهای هتروکروماتیک با تکنیکهای باندینگ C و همچنین با روش های مختلف رنگ آمیزی فلوروکروم مشخص میشود و با مفهوم هتروکروماتین کلاسیک تعریف شده، مطابقت دارد. نواحی هتروکروماتیکی کروموزومها به طور معمول در کل اینترفاز متراکم باقی میمانند و عموماً به عنوان بلوکهایی در اطراف سانترومرها یافت میشوند، اما گاهی اوقات در نواحی انتهایی با بینابینی کروموزومها قرار میگیرند.
- باندهای یوکروماتین الگویی از باندهای متناوب رنگ آمیزی مثبت و منفی (یا فلورسنت) را در طول کل کروموزومها تشکیل میدهند و توسطG-banding ، R-banding، Q-banding و «فلوکرومها» (Fluorochromes) خاص نشان داده میشوند. یوکروماتینها نسبت به هتروکروماتین درون هسته بسیار کمتر متراکم بوده و از نظر ژن غنی هستند.
- نواحی سازمان دهنده هستهای بخشهایی از کروموزومهایی هستند که حاوی ژنهای RNA ریبوزومی بوده و باعث بوجود آمدن هستههای اینترفاز می شوند. آنها را میتوان با نقره رنگ آمیزی کرد (رنگ آمیزی Ag-NOR).
- کینه توکور ساختارهای سانترومری هستند که از طریق آنها کروموزومهای میتوزی و میوزی به میکروتوبولهای دوک تقسیم وصل میشوند و به طور کلی با استفاده از سرمهای خود ایمنی CREST برچسب گذاری میشوند.
باند R زمینه روشن
باند R تقریباً معکوس باندهای G است (R مخفف کلمه reverse معکوس است). نواحی تاریک یوکروماتینی و نواحی روشن از نوع هتروکروماتین هستند.
رنگآمیزی DNA
برای شناخت چگونگی ایجاد باندها در روش باندینگ و نحوه رنگآمیزی ساختارهای کروموزومی در درجه اول باید به درک چگونگی اتصال رنگها به DNA پرداخت. روشهای مختلفی برای اتصال رنگهای مختلف به DNA وجود دارد. برخی از رنگهای مورد استفاده در این روشها برای DNA اختصاصی هستند و تنها به رشتههای DNA متصل میشوند و اتصالی با پروتئینهای کروموزومی ندارند.
رنگهای باندینگ کروموزومها به چند روش مختلف به DNA متصل میشوند. یکی از این روشها اتصال و برهمکنشهای رنگها با DNA از طریق شیارهای بزرگ و کوچک موجود در ساختار دو رشتهای DNA است. این برهمکنشها میتواند از جنس پیوندهای شیمیایی مختلف یا کووالانسی باشد. در برخی موارد پیوندهای هیدروژنی و برهمکنشهای الکترواستاتیک میتواند باعث اتصال رنگها به ساختار DNA شوند.
در برخی از موارد نیز رنگهای مورد استفاده در روشهای باندینگ ممکن است چندین گروه عاملی داشته باشند و از این رو، از جایگاههای مختلف به ساختار کروموزومها میتوانند متصل شوند.